
Млекопитающие — наиболее высокоорганизованные позвоночные животные, освоившие все среды обитания и приспособившиеся к самым различным способам передвижения. Размеры у представителей класса сильно варьируют: от 3,5 см у карликовой белозубки до 33 м у голубого кита. Среди млекопитающих наиболее распространены наземные формы, представлены подземные, водные и летающие.
Среди наземных форм выделяют стопоходящих (медведи), пальцеходящих (большинство видов), пальцестопоходящих (некоторые зайцеобразные, виверровые) и фалангоходящих (копытные). Стопохождение обеспечивает относительно медленное передвижение. Пальцехождение и особенно фалангохождение, связанное с редукцией части пальцев, дают возможность быстрого бега.
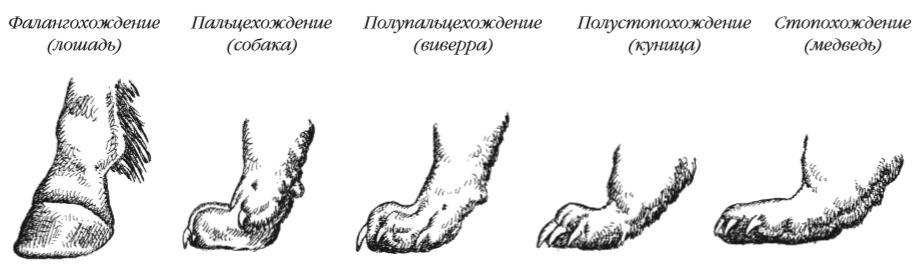
Основную локомоторную функцию у китообразных выполняет хвост с горизонтально расположенными и бесскелетными плавниками. Модифицированные конечности (ласты) имеют ластоногие и сирены, а для кротовых характерны копательные лапы.
Волосы и другие производные эпидермиса
Волосяной покров, свойственный для млекопитающих, является роговым образованием эпидермиса. Основное назначение волосяного покрова — термоизоляция. Волосы подразделяются на вибриссы, выполняющие осязательную функцию, на направляющие, остевые и пуховые. Примером видоизмененных волос служат щетина и иглы. Для млекопитающих характерна сезонная смена волосяного покрова (линька).
К роговым образованиям эпидермиса относятся чешуи, когти, рога (у полорогих парнокопытных), нёбные роговые валики, мозоли и челюстные чехлы.
Чешуи в качестве основного покрова имеют ящеры и броненосцы. (У броненосцев чешуи подстилаются костными пластинками.) Чешуйчатый покров на хвосте в той или иной степени встречается у многих млекопитающих (бобр, выхухоль, мышевидные грызуны, сумчатые и другие). Рудименты чешуй находятся в области спинного плавника у некоторых китообразных (морские свиньи, гринды).
Когти и их модификации (ногти, копыта) предохраняют конечные фаланги пальцев млекопитающих от повреждений, используются при ловле добычи, рытье нор и лазании. Коготь состоит из плотной когтевой пластинки, прикрывающей последнюю фалангу сверху и с боков, и рыхлой подошвенной пластинки. У приматов когтевая пластинка уплощена и прикрывает фалангу только сверху (ноготь), а подошвенная пластинка сильно редуцирована. Копыто образовано мощно развитыми когтевой и подошвенной пластинками, а также ороговевающей подушечкой пальца, которые со всех сторон охватывают фалангу.
К эпидермальному образованию относится рог носорога. Он представляет собой сплошную роговую массу из склеенных между собой нитей, образующую прочный конус над носовыми костями. Рога полорогих копытных — полые роговые чехлы, сидящие на костных стержнях, которые очень рано срастаются с лобными костями. У полорогих рога растут медленно в своем основании и сохраняются на всю жизнь. (Рога оленей имеют другое происхождение. Они развиваются из кориума и состоят из костного вещества.)
Нёбные роговые образования представлены у китообразных в виде пластин (китовый «ус») или нёбных валиков (у сирен). Челюстные роговые чехлы у однопроходных (утконос, ехидна) функционально заменяют отсутствующие зубы.
Кожные железы
Кожные железы образуются из эпидермальных зачатков, погруженных в толщу кориума (дермы). Характерны два основных типа желез — трубчатые и гроздевидные. К первым относятся потовые железы, имеющие вид спирально изогнутой трубки с концом, свернутым в клубочек. Их протоки открываются на поверхность кожи или в верхней части волосяной сумки. Эпидермальные клетки этих желез выделяют пот, состоящий из воды, в которой растворены мочевина, некоторые соли и жирные кислоты. Хотя вместе с потом выделяется часть продуктов распада, основное назначение потовых желез — терморегуляция. Выделяющийся при перегреве животного пот испаряется и охлаждает тело. Потовые железы обильно представлены у приматов и копытных, слабо развиты у волчьих, кошачьих, грызунов и зайцеобразных. У китообразных, ленивцев и ящеров они полностью отсутствуют. Эти животные используют иные формы терморегуляции.
Сальные железы имеют гроздевидное строение и открываются в волосяные сумки. Стенки желез образованы многослойным эпителием. Клетки эпителия претерпевают жировое перерождение и образуют жирный секрет, который обеспечивает эластичность кожи и волос.
Пахучие железы возникают за счет видоизменения потовых или сальных желез (или их комбинаций) и выделяют пахучий секрет. К ним относятся анальные железы куньих, мускусные железы кабарги, бобров и выхухоли. У парнокопытных (олени, антилопы, горные козлы и бараны) развиты предглазничные и межкопытные пахучие железы (см. рисунок). Пахучий секрет используется для мечения территории и видового опознавания.
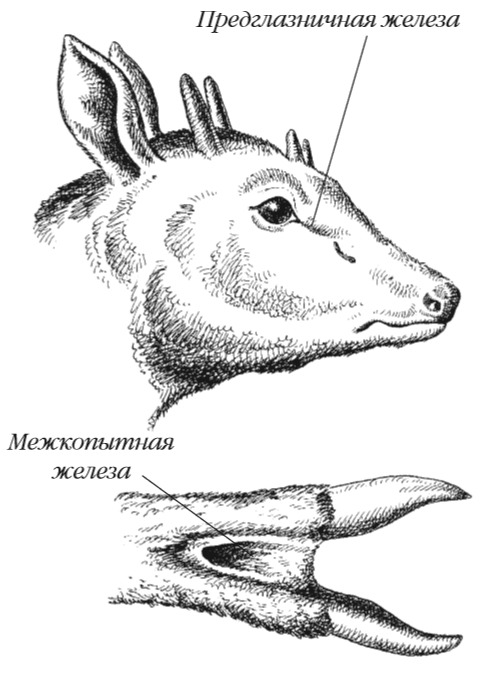
У некоторых животных секрет анальных желез, имеющий резкий неприятный запах, применяется для пассивной защиты (американские скунсы, некоторые хорьки). Совокупность запахов пахучих, сальных и потовых желез позволяет животным опознавать особей своего вида, облегчает встречу самцов и самок. Обладая высоко развитым обонянием, млекопитающие прекрасно ориентируются по пахучим «меткам», различая «своего» и «чужого».
Млечные железы образованы модификациями потовых желез. У однопроходных они сохраняют трубчатое строение и представлены железистыми полями (у утконоса — на брюхе, у ехидны — в выводной сумке). Протоки желез у этих животных открываются в волосяные сумки, и детеныши слизывают выступившие капельки молока. Остальные млекопитающие имеют молочные железы гроздевидного строения, протоки которых открываются на сосках. Соски могут располагаться двумя рядами от передних конечностей до паховой области (насекомоядные, хищные, грызуны). У приматов, слонов, рукокрылых и сирен сохраняется только грудная пара сосков. Для большинства копытных характерно слияние молочных желез в расположенное в паховой области вымя, имеющее два или четыре соска. В целом число сосков у млекопитающих варьирует от 2 до 24 и зависит от количества рождаемых детенышей. Наибольшего развития млечные железы достигают в период беременности и выкармливания молодняка.
Размножение и забота о потомстве
Половой диморфизм у большинства млекопитающих выражен достаточно слабо — самцы несколько крупнее самок, отличаются более грубым волосяным покровом. Резкие половые отличия характерны для кашалотов, ушастых тюленей и некоторых приматов. У них самцы значительно превосходят по размерам самок. Напротив, самки крупнее самцов у усатых китов. Клыки, бивни, рога, грива более развиты у самцов или свойственны только им (львы, ушастые тюлени, олени). Половой диморфизм может отчетливо проявляться в окраске, как это иногда имеет место у некоторых копытных. Вторичные половые признаки облегчают взаимную встречу партнеров, способствуют синхронизации половых циклов у самцов и самок.
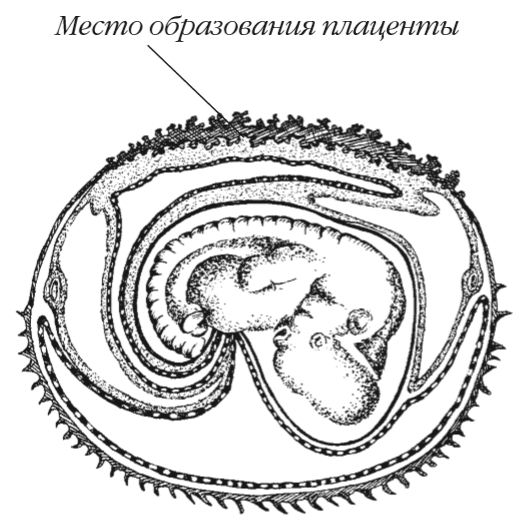
Для млекопитающих свойственно внутриутробное развитие зародышей за счет питательных веществ, получаемых от матери через особый орган — плаценту.
(Однопроходные — утконос и ехидна — откладывают богатые желтком яйца.) Примитивная плацента характерна для зародышей некоторых видов сумчатых.
Она образуется путем срастания наружной стенки аллантоиса с серозой, но не имеет ворсинок, проникающих в толщу стенок матки. Питание зародыша при подобном типе плаценты осуществляется за счет желтка и секреторных выделений стенки матки.
Настоящая плацента, свойственная для большинства млекопитающих, возникает при срастании участков серозы и наружной стенки аллантоис и сопровождается образованием выростов (ворсинок).
Выросты внедряются в стенку матки, которая в этом месте разрыхляется и образует материнскую часть плаценты. Кровеносные сосуды зародыша приходят в более тесное (по сравнению с примитивной плацентой) соприкосновение с сосудами материнского организма. Это улучшает его питание за счет веществ материнской крови (гемотрофное питание).
Продолжительность беременности у сумчатых невелика. Например, для опоссума она составляет 12 дней. Детеныши рождаются очень маленькие (у опоссума — меньше пчелы) и недоразвитые. Они донашиваются в сумке, в полость которой открывается сосок. Молоко впрыскивается в рот детеныша сокращением специальной мышцы, а сам детеныш висит на разбухшем соске. У плацентарных млекопитающих детеныши рождаются более развитые. Во всяком случае, они способны самостоятельно сосать молоко, а у некоторых видов (копытные, киты и другие) способны активно передвигаться вслед за матерью.
Количество детенышей, приносимых самкой, варьирует от 1 (китообразные) до 15-20 (некоторые сумчатые и грызуны). Продолжительность выкармливания молоком весьма изменчива — от 10-20 дней (мелкие виды грызунов) до 2 лет (моржи).
К концу лактационного периода родители переводят своих детенышей на взрослый корм. Они приводят их на места кормежки или приносят корм к логову. Хищники часто используют еще живую добычу, что позволяет малышам вырабатывать «охотничьи» навыки, необходимые для будущей самостоятельной жизни.
В случае частого беспокойства или истощения запасов корма на данной территории родители переносят своих детенышей в другое место. Чаще всего это делает самка. У некоторых видов млекопитающих самки более или менее постоянно носят детенышей на себе.
При опасности, грозящей гибелью потомству, мать пытается отвлечь внимание врага. Так поступают многие хищники, даже олени и лоси, детеныши которых в это время затаиваются. Стадо овцебыков при нападении хищников образует вокруг телят защитное кольцо. Самки моржей и китов не покидают своих детенышей, что бы им ни грозило.
Связь между самкой (родителями) и потомством может сохраняться некоторое время и после завершения выкармливания. Этот важный период обучения позволяет развивать и накапливать популяционно-видовые навыки.
Происхождение млекопитающих
Предками млекопитающих считают древнейших пресмыкающихся. К ним принадлежала жившая в пермском периоде и триасе группа, объединяемая в подкласс звероподобных (Theromorpha). Особенно близок к млекопитающим был отряд зверозубых (Theriodonta), представители которого имели дифференцированную зубную систему, сходное с млекопитающими деление позвоночного столба, строение поясов конечностей и самих конечностей.
Некоторые из зверозубых отличались достаточно крупными размерами — например, иностранцевия (Inostrancevia alexandrovi), известная из пермских отложений и достигавшая в длину 3 м. Однако предками млекопитающих скорее всего явились мелкие зверозубые, близкие к иктидозаврам (высшим териодонтам) из верхнего триаса.
Остатки древнейших млекопитающих известны из юрских отложений. Наиболее примитивными из них были многобугорчатые (Multituberculata), имевшие большое число бугорков на коренных зубах. Зубная система этих животных напоминала особенности зубов у грызунов. Клыки отсутствовали, а резцы отделялись от коренных зубов выраженной диастемой. Mногобугорчатые, как предполагают, были растительноядными, и, возможно, яйцекладущими формами. В нижнем эоцене все они вымерли.
Жившие в юрском периоде другие древнейшие ветви — трехконусозубые (Triconodontia), симметродонты (Symmetrodontia) и трехбугорчатые (Tritubetculata) — прекратили свое существование в конце мелового периода. Наибольшим разнообразием отличались трехбугорчатые, или пантотерии (Panthotheria). К ним относились мелкие зверьки, напоминающие современных насекомоядных. Коренные зубы у них несли по три тупых бугорка, расположенных асимметричным треугольником (см. рисунок). Именно пантотериев считают вероятными предками сумчатых и плацентарных.
Сумчатые возникли в конце мелового периода и были представлены мелкими формами (близкие к опоссумам). Центрами возникновения сумчатых считают Евразию и Северную Америку. Не выдержав конкуренции с заселившими эти континенты плацентарными, большинство сумчатых вымерло до конца третичного периода. Плацентарные не проникли в Австралию, где сумчатым удалось сохраниться и в известной мере процветать. Центром возникновения плацентарных в меловом периоде послужила Азия, откуда шло их расселение в разных направлениях. Древнейшие плацентарные принадлежали к насекомоядным, послужившим предковой формой для возникновения других отрядов. Что касается происхождения однопроходных, то наиболее вероятным считается их родственная связь с древнейшими трехконусозубыми (Hconodontia), от которых они обособились достаточно рано. В юрском периоде сформировалось 6 отрядов млекопитающих, а в палеоцене — не менее 16, из которых Однопроходные (Monotremata), Сумчатые (Marsupialia), Насекомоядные (Insectivora), Шерстокрылы (Dermoptera), Неполнозубые (Edentata), Приматы (Primates), Грызуны (Rodentia), Хищные (Carnivora) и Зайцеобразные (Lagomorpha) сохранились до настоящего времени.
С эоцена в Северной Америке и Евразии развивалась ветвь титанотериев (Titanotheria) — гигантских животных с раздвоенным рогом на носовой части черепа (см. рисунок). Свое существование эта ветвь прекратила в олигоцене.

Среди вымерших гигантских безрогих носорогов выделялся индрикотерий (Indricotherium), достигающий в длину 7 м, а в высоту — 5 м. В конце плейстоцена жил шерстистый носорог (Coelodonta antiquitatis) с огромным рогом. В эоцене появились древнейшие Хоботные (Proboscidea) — мастодонт (Gomphotherium), а в плейстоцене — мамонт (Mammuthus primigenius).
Становление класса млекопитающих осуществлялось достаточно медленно. У звероподобных пресмыкающихся наиболее развитым отделом головного мозга был мозжечок. В процессе эволюции у млекопитающих происходило постоянное увеличение переднего мозга. Последовательное накопление адаптаций (четырехкамерное сердце и теплокровность; живорождение и выкармливание детенышей молоком; развитие центральной нервной системы и органов чувств) обеспечило млекопитающим успешную конкуренцию с пресмыкающимися. Интенсивное вымирание динозавров в конце мезозоя позволило млекопитающим наряду с птицами занять господствующее положение.
Класс млекопитающих (Mammalia) включает 19 современных отрядов, объединяющих около 4000 видов (по разным данным — от 3700 до 4237).


